未来発生進化学: 研究室が目指すもの
進化、というと、「鳥の羽は飛ぶために進化した」とか、「白熊が白いのは、背後の雪や氷の色に溶け込むのに適している」など、目的論的な議論を思い浮かべる人が多いかもしれません。確かにこのような議論は、時として面白い小話を提供してくれます。しかし、前提となる進化的原理についての仮説が変われば、解析法が変わり、結論も大きく変わってくるでしょう。例えば、「進化は変異と自然淘汰によって起こる」と考えるか、「進化は環境によって誘導された形質によっても起こる」と考えるか、それによって、出来上がる「小話」は違ったものになるのです。
この研究室が目指すのは、前提となる進化的仮説のうえに現在の生物界を理解するのではなく、現在の動物を分子生物学的に研究することで、進化における形態形成の変化の法則を明らかにしていくことです。進化生物学の分野では、これまで「自然淘汰がどのような形質を選んだか」についてたくさんの研究が行われてきました。多くの研究者は、「ランダムに生じた形質を自然淘汰が淘汰することによって現在の生物のかたちが生まれた」と考えていますが、その逆の仮説、つまり「環境が特定の形質を生み出した」という仮説についてはほとんど研究が進んできませんでした。
半世紀以上前、英国の著名な発生生物学者であるConrad Hal Waddingtonは、「ゲノム中に隠れて普段は形質として現れないcryptic genetic variationが、それぞれの環境に応じて淘汰されて、環境に応答した形質が生まれる」とするgenetic assimilation仮説を提唱しました。さらにその後、米国の著名な進化生物学者Mary Jane Eberhardは、「環境に応答した生物の可塑性が、新たな表現型の源泉となっている」という仮説を提唱しました。どちらの仮説にも、実験的なデータがありますが、実際の進化で果たした役割についてはほとんど明らかになっていません。
脊椎動物は、大きな脳を含む中枢神経系や顔面の構造、運動に適した尾や運動器官を発達させ、この地球上で大きく分布域を拡大しました。こうした脊椎動物にみられる特徴は、どこから生まれてどうして進化したのでしょうか? 本研究室では、脊椎動物が無脊椎動物の祖先から進化した際に、地球における環境変動が脊椎動物の特徴の進化に与えた影響を明らかにするため、脊索動物の発生にかかわる遺伝子ネットワークの熱ストレスに対する可塑性を明らかにし、発生システムの進化と環境とのかかわりを明らかにしようとしています。
また、生物の初期発生は生物の特徴を決定するカギとなりますが、この段階では、母親から与えられた母性RNAが発生を支配していますので、環境によって母親から与えられる母性RNAがどのように変化するかということにも着目して研究を行っています。この取り組みのひとつとして、東京海洋大学の吉崎悟朗研究室が開発した代理親魚法と呼ばれる手法を用い、母親をすり替えた場合に生殖細胞の母性RNAやその後の発生に起こる変化を明らかにする研究も行っています。
こうした新たな進化的原理を明らかにしていくことにより、現在の生物の生物の研究によって過去を語るだけでなく、未来に向かって生物がどのように変化していくのか、また、われわれの暮らしや社会のあり方によって、その未来がどう変わっていく可能性があるかを考える基盤を提供していくことが、本研究室の目標です。100年先の未来の人類が、自分たちの生き方を考える礎となる科学的原理を提供したい。そういう思いで、今日も研究に励んでいます。
1. 発生緩衝の分子メカニズム
生命現象はすべて熱力学的に解釈することが出来る、という研究者が多くいます。しかしそれは本当でしょうか。そうだとするとそれは面白いけれど、そうでないとしたらそれはもっと面白い。生物には、生物にしかない法則も持っているということになるからです。生物にしかない法則かもしれない、と研究をすすめているもののひとつが、発生緩衝です。発生緩衝とは、環境やゲノムの変異によらず、発生が安定的に進むことを可能にする仕組みのことです。発生緩衝は、ある表現型が発現するかどうかを左右する、進化における要です。発生における表現型として発現するか否かがどのようにして決定しているかを明らかにすることは、生物の進化の法則を明らかにすることにつながります。
発生緩衝のしくみとしては、これまでシャペロンの一種Hsp90やネットワーク理論などの研究が行われてきました。当研究室の活動では、発生緩衝が母親由来であること、つまり母親が子の発生の安定性を決めていることを明らかにしました(Sato et al. 2015)。そして、温度環境に比較的強い種と弱い種のカタユウレイボヤにおけるトランスクリプトームの比較解析から、ゲノムに存在すると考えられている全てのシャペロン分子について発生緩衝との関連を研究し、これまで考えられてきた細胞質内に存在するHsp90ではなく、これまで発生との関係において全く注目されてこなかった、小胞体に関連したシャペロンが重要であることを発見しました(Sato et al. 2015)。この結果は、カタユウレイボヤだけでなく、線虫を用いた実験でも検証され、動物界で広く保存されいることが示唆されました(Hughes et al. 2019)。さらに、これらの小胞体関連シャペロンには、新しく翻訳されたアミノ酸の鎖を正しく折りたたむだけでなく、余分なタンパク質や壊れたタンパク質を壊す役割を担うシャペロンも含まれていることを明らかにしました(Sato et al. 2015; Hughes et al. 2019; Sato 2018)。タンパク質の合成だけでなく、タンパク質の分解も重要であることを提案しました(Hughes et al. 2019; Sato 2018)。この考え方は、生物時計など、他の幅広い発生モデルで検証され、受け入れられつつあります。
当研究室ではさらに、発生緩衝の遺伝子ネットワークの側面からの理解も進めています。なぜなら、ネットワークとして理解することで、その状態を数式化し、数学的に予測可能なものにしたいと考えているからです。これまでの当研究室での研究では、熱ストレスに比較的強い種と弱い種のカタユウレイボヤのトランスクリプトームデータ解析から、発生緩衝の分子ネットワークは、発生のネットワークと驚くほど弱いつながりしかないことが明らかになりました。発生緩衝のネットワークは、形態形成に重要なシグナリング分子とつながっていることも明らかになり、シグナリング分子の安定性が発生緩衝において非常に重要であることが示唆されました。現在は、発生緩衝が弱い場合に発生ネットワークのどの部分が壊れてしまうのかを明らかにし、それによって、発生と環境との関係を数理的に理解する取組みを行っています。
2. 母性mRNAの由来と、その編集の研究
当研究室では、発生緩衝の度合いが母親によって決まることを明らかにしてきました(Sato et al. 2015)。そこで当研究室が次に着目したのが、母親の環境によって卵の遺伝情報が変化する可能性です。例えば、近年の研究では、分子量の小さなRNAが、親の環境や栄養状態によって配偶子に輸送され、配偶子のエピジェネティックな状態を変化させることが報告されています。母性RNAはDarwinの時代からこれまで、母親由来ゲノムの転写物であると盲目に信じられてきましたが、本当にそうでしょうか。母親の体細胞由来のmRNAは、卵に存在しないのでしょうか。もし存在する場合、それはどのように輸送され、どのような役割を果たしているのでしょうか。こうした可能性について、異なった温度環境に適応しているカタユウレイボヤ(図1)の2種や、線虫(図2)、また魚類(図3)をモデルに研究を進めています。これまで、生殖細胞を、もともとの親の体内以外の場所、つまり他の動物の生殖巣や研究室内の培養細胞の環境で生産するという技術の開発が急速に進められていますが、例えば、ヒトの生殖細胞をマウスの生殖巣由来の培養細胞を使ってつくろうとしてもうまくいきません。このような異種間での生殖細胞形成が、組み合わせによってうまくいかないというのも、体細胞-生殖細胞間のコミュニケーションにおける問題を示唆していることが考えられます。体細胞-生殖細胞間のコミュニケーションを解明する取り組みは、こうした生殖細胞の増殖技術にも応用される可能性があります。
-

- 図1

-
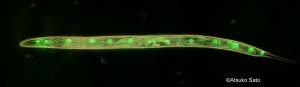
- 図2
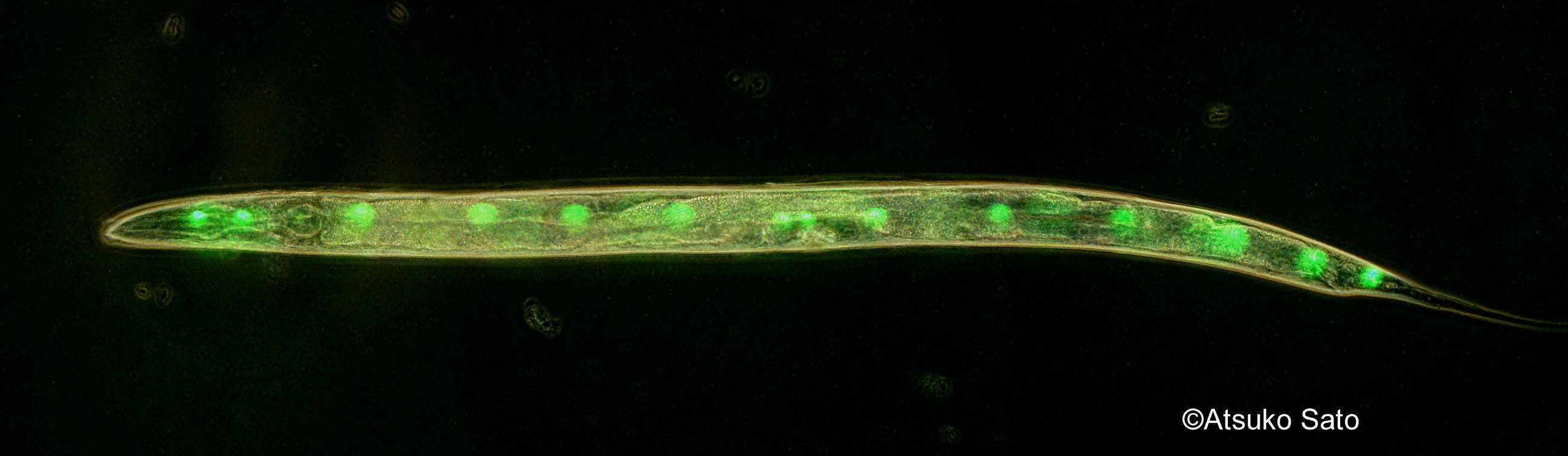
-

- 図3

3. 環境が子孫に与えるばらつきの研究
Charles Darwinは、環境の影響が、進化の根源となるバリエーションを生む鍵となっていることに気づいていました。そしてこれまで100年以上にわたって、どのようにしてバリエーション(つまりばらつき)が生まれるのかについて、様々な仮説が立てられてきました。母親の環境が子の発生や発育に大切であることは以前からも言われてきたことですが、進化の観点から集団としてみたときに、母親の環境はどのような影響を及ぼしているのでしょうか。変動する環境下では多様な集団が生まれ、その一部が生き残るという戦略が採られているという説や、発生緩衝が強い集団では、表現型にならない変異が蓄積し、環境が変動したときにのみそれらが表現型として現れることでバリエーションを高めているという説など、これまで様々な説が提唱されています。当研究室では、発生緩衝度合いが顕著に異なるカタユウレイボヤの姉妹種において、継代飼育を行い、シングルセルレベルで卵に観察されるばらつきを解析することにより、この問題にこたえる国際共同研究を、John Bishop (Marine Biological Association of the UK) やManuela Truebano (Plymouth University)らとともにすすめています(Sato, 2020; Sato et al. 2024)。Waddingtonが提唱したGenetic Assimilation 仮説について、脊索動物の進化にどのような影響を与えたかについて、エピゲノムの観点からの研究も行っています。
4. 可塑性と生物多様性との関係の解明
生物の可塑性は、生物学者を魅了し続けてきました。進化は、変異と淘汰だけで説明が出来る、とする説に対し、これまで多くの研究者が、環境によって引き出される可塑性こそが、進化の駆動力になっているという仮説を提唱してきました。近年では、エピジェネティクスなどの新しい分子レベルでの仕組みも明らかになりつつあり、Extended Evolutionary Synthesisという、進化生物学を二分する大きな議論を巻き起こしています。しかし、実際にこれらの仮説のどちらが正しいかを検証することは非常に難しく、議論は今も続いています。その大きな理由のひとつが、可塑性によって現れる複数の表現型のうち、ひとつだけが集団内に固定する分子レベルでのメカニズムが明らかになっていないことです。集団内に固定するメカニズムは、DNAの変化によるものなのか、それともエピジェネティックに説明できるのか。
この問題に、当研究室は、日本に馴染みの深いサザエの例を利用して取り組みたいと考えています。サザエは、日本を中心とする東アジアでしか見られない種です。一種の中に、棘を持つものと持たないものが現れ、さらには、一個体の中でも、棘を作る時期や作らない時期がある個体もあります(図4)。このような大きな可塑性を示す生物は、10万種あるといわれる貝類の中でも非常にわずかであり、サザエほど非常に柔軟性に富んだ可塑性を示す貝類は他にないと思われます。一方、サザエに非常に近縁なチョウセンサザエでは、まったく棘が現れません。一方の、カタベガイなどでは、棘が常に現れ、棘が無い個体はありません。サザエを用いて棘を形成する生態レベルおよび分子レベルでのメカニズムを明らかにすることにより、環境による刺激が進化的新規性の根源となっているとする、phenocopyやgenetic assimilationなどの歴史的な議論に新たな証拠を与えることが出来るのではないかと考えています。サザエの表現型を切り替えている要因やその仕組みについては、波の強さや環境の影響であるとする生態学的な仮説がありますが、未だ明らかではありません。当研究室では、この可塑性の原因を明らかにし、数学的に予測することを目指し、貝殻の形態と数理モデル、さらに解析の自動化を組み合わせた国際共同研究を、Derek Moulton (University of Oxford, Mathematical Institute)およびNathanael Aubert-Kato (お茶の水女子大学 情報科学科)とともにすすめ、分子レベルでの化学反応について、貝殻の形態からどのような予測が可能か、また、環境によって固有の数理モデルが得られるのかについて、研究を行いました(Moulton et al. 2024)。
サザエは日本に大変馴染みが深く、数年前までは、ある水産技術センターの職員さんが、「日本は魚がとれなくなってもサザエを食べて生きていける」などと言っていたほどでしたが、最近になって、千葉県、神奈川県の一部、紀伊半島、そして高知県など、全国各地から、サザエが激減しているという情報が入っています。生態学的な正確なデータはまだ得られていませんが、えさとなる海藻群落が激減していることと関連しているのではないかと考えられています。日本近海の海の資源を守るよう、早急な対策が必要です。
 図4
図4
5.翼鰓類の研究
半索動物は、われわれ脊椎動物の起源とその進化について研究する上で非常に重要な動物です。半索動物に見られる2属のうち、腸鰓類(図5)は比較的よく研究されていますが、翼鰓類(図6)についての生物学的知見は非常に限られています。イギリスで採集されるRhabdopleura compactaを題材とし、生態や発生、また進化発生学的な研究を行っています。
-

- 図5

-

- 図6
